




Nous tenons à remercier spécialement tous les vétérinaires de la SFEROV (Société Française d'Etude et de Recherche en Ophtalmologie Vétérinaire) qui nous ont beaucoup aidé dans l'élaboration de ces chapitres que nous allons encore compléter.
Merci beaucoup au Pr Francis Lescure, au Pr Raynier, à Valérie Meunier et à tous leurs collègues.
La physiologie et la pathologie comparées sont deux des chapitres les plus passionnants de la biologie et de la médecine. L'une et l'autre s'élèvent au dessus du caractère purement utilitaire de la connaissance. Elles incitent à la réflexion sur le pourquoi de tant de différences. Quand la fonction reste la même et que le but à atteindre est commun. La vision et sa première composante, la sensation visuelle, sont un des exemples typiques de ce genre de réflexion.
0n peut répondre d'emblée que le besoin n'est pas le même selon l'espèce, en particulier selon le milieu dans lequel elle vit, la rapidité avec laquelle elle se déplace, la nourriture qu'elle doit chasser et saisir, la vigilance dont elle doit faire preuve pour échapper elle-même aux prédateurs, tout cela est vrai et satisfaisant pour l'esprit, mais seulement d'une façon partielle.
Pour s'en convaincre, il suffit d'étudier la performance de l'oeil chez les différentes espèces de vertébrés. Mais que faut-il entendre par performance oculaire ?
La performance oculaire est le degré de perfection de l'image formée sur la rétine à partir de l'objet qui envoie des rayons lumineux sur l'oeil, permettant à l'animal d'apprécier la luminosité, la forme, les dimensions, la position dans l'espace et éventuellement couleurs et mouvements. En d'autres termes, la performance oculaire c'est la qualité de la vision ramenée à l'étage oculaire, étant bien entendu que dans la vision définitive, interviennent largement la qualité des voles optiques et l'aptitude des centres visuels.
Cette performance peut être scindée en plusieurs éléments, essentiellement quatre :
- La netteté, telle qu'à un point de l'objet corresponde un point de l'image.
- La résolution qui est la plus petite distance séparant deux points objets capables de donner deux points images distincts.
- Le contraste, reproduction de la différence d'éclairement des surfaces distinctes et éventuellement de leurs couleurs.
- Les mouvements de tout ou partie de l'objet.
Ces quatre éléments vont être étudiés séparément en parcourant l'échelle animale.
A/ La netteté de l'image
La netteté est la résultante de trois processus :
La réfraction statique, fruit de la convergence de la cornée et du cristallin.
La réfraction dynamique ou accommodation qui modifie la convergence selon la position de l'objet par rapport à l'oeil.
La correction de l'aberration marginale par lejeu du diaphragme irien.
1/ La réfraction statique
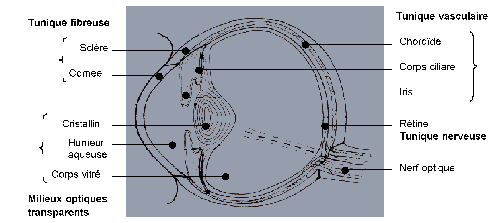
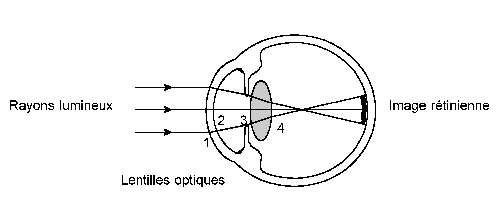
Elle est telle que, dans l'oeil emmétrope, les objets situés à plus de 5 mètres donnent une image exactement sur la rétine, autre ment dit la rétine se trouve au foyer du dioptre oculaire. Il n'est pas le lieu, ici, de reconstruire l'oeil théorique et d'étudier le cheminement des rayons lumineux dans les différentes structures de l'oeil. Supposant cela acquis, ne considérons que les différences spécifiques.
La première différence oppose les animaux terrestres aux animaux aquatiques car chez ces derniers, par suite de la similitude de réfraction eau-cornée, la cornée n'intervient pas. La cristallin est donc seul responsable de la réfraction statique. Pour assumer cette fonction, le cristallin est très globuleux et parfois même sphérique ; en outre, il se situe le plus loin possible de la rétine, c'est-à-dire presque au contact de la cornée ; la chambre antérieure est alors presque virtuelle.
Le problème est plus complexe chez les amphibiens et les poissons partiellement immergés tels que l'anableps. Ce dernier, improprement qualifié de « tétrophtalme », possède une pupille divisée en deux par des expansions latérales de l'iris : une pupille ventrale et une dorsale. Le cristallin est ovale à grand axe oblique, de telle sorte que les rayons lumineux venant d'objets immergés passent par la pupille ventrale, traversent le cristallin suivant le plus grand axe et convergent sur la rétine dorsale.

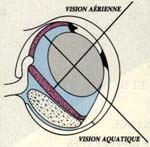
A l'inverse, les rayons venant d'objets aériens sont réfractés par la cornée puis par le petit axe du cristallin et convergent sur la rétine ventrale.
L'étude de la réfraction statique chez les vertébrés terrestres n'a pas encore bénéficié de tous les moyens techniques dont on dispose aujourd'hui. D'une façon générale, on admet que les espèces pourvues de petits yeux sont hypermétropes, tandis que les gros yeux sont myopes.
A la vérité, il n'y a pas eu d'étude très poussée sur ce sujet en raison de l'absence d'intérêt pratique sauf peut-être pour le cheval et notamment le cheval d'obstacle. Chez celui-ci, la mesure de la réfraction a été faite par rétinoscopie et par skiascopie.
On a pu mettre en évidence, à côté de chevaux parfaitement emmétropes, la présence de myopes et d'hypermétropes. Ces examens cependant n'ont pas tendance à se généraliser en l'absence de sanction thérapeutique.
Il en ira différemment le jour où l'on pourra pratiquer chez le cheval une chirurgie réfractive correcte.
Les affirmations générales concernant le chien ne peuvent relever que de la plus grande fantaisie. Il suffit de regarder l'oeil des différentes races pour comprendre le danger de toute généralisation. On peut affirmer qu'il existe dans toutes les races des emmétropes et des amétropes qui mériteraient d'être corrigés.
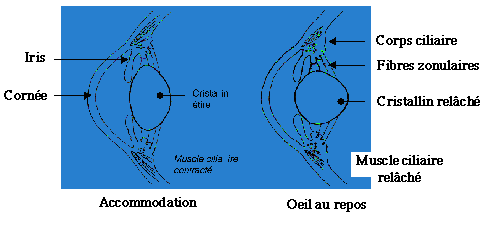
2/ La réfraction dynamique ou accommodation
Elle permet à l'animal de diminuer ou d'augmenter la convergence de l'oeil, par un phénomène actif ou par un phénomène passif.
=> La diminution de convergence
Elle est surtout observée chez les poissons téléostéens par un mouvement de recul ou de retrait du cristallin. Le retrait s'effectue par contraction du muscle Retractor lentis. Il faut noter que le retrait par mouvement latéral n'est jamais complet et l'ouverture pupillaire n'est que partiellement aphaque.
=> L'augmentation de convergence
Elle est beaucoup plus fréquente que la première et permet la vision des objets rapprochés.
Elle peut être passive ou active.
* Réfraction passive :
La rétine en pente
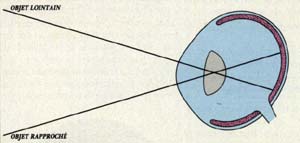
La distance de la rétine à la cristalloïde postérieure varie selon le méridien vertical : plus éloignée dans la région supérieure pour la vision des objets rapprochés ; plus rapprochée en bas pour la vision des objets lointains. Cette disposition s'observe chez les sélaciens (raie, requin) et chez le cheval. Ce dispositif est plus facilement admis pour les sélaciens que pour le cheval. Chez lespoissons, les rayons incidents traversent la cornée sans être déviés ; le cristallin sphérique dévie de la même manière tous les rayons incidents, tous convergent sur une même partie de la rétine et la rampe joue bien son rôle optique .
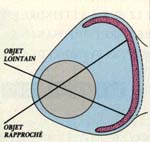
Chez le cheval au contraire, la cornée dévie les rayons incidents et le cristallin se comporte comme un prisme, ramenant les rayons en direction de l'arca striaeformis, bande horizontale, à la limite inférieure du tapis, de plus grande sensibilité rétinienne .
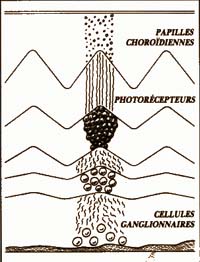
Les papilles choroïdiennes (fig. 4)
Chez les chiroptères (grandes chauves-souris frugivores et roussettes). Elles donnent à la rétine un aspect ondulé et aux photorécepteurs un éloignement plus ou moins grand selon leur position sur ou dans un pli.
* Réfraction active :
Par modification de. la courbure cornéenne, elle intervient chez les oiseaux. Le muscle ciliaire est divisé en deux portions : l'une postérieure peut modifier la courbure de la cornée, c'est le muscle de Crampton, l'autre antérieure modifie le cristallin, c'est le muscle de Brücke.
Par modification de la courbure du cristallin
Augmentation de la courbure des cristalloïdes par relâchement de la zonule après contraction du muscle ciliaire. C'est le phénomène que l'on observe chez les mammifères ;
Transformation sphérique d'un cristallin mou par striction exercée par le sphincter irien. Cela permet une accommodation maximale chez les chéloniens (tortue) et chez les oiseaux, notamment lesrapaces.
Par changement de position du cristallin
Tiré vers l'avant, chez les amphibiens (grenouilles) ou poussé vers l'avant par action du vitré chez les ophidiens (serpents).
Poussé vers l'arrière par pression sur la cornée du muscle extra-oculaire cornéen chez les cyclostomes (lamproies) et les lacertiliens (lézards) ;
Tiré en arrière par le muscle rétracteur, campanule de Haller des téléostéens.
3/ Le diaphragme irien et la pupille
Le rôle du diaphragme irien est double:
éliminer les rayons marginaux et protéger la rétine d'un excès de luminosité. Les animaux nocturnes ont une pupille ronde et large.

Les nocturnes qui émergent le jour ont une pupille en fente, qui, beaucoup mieux qu'une pupille ronde, peut s'obturer complètement.
Tous les oiseaux ont une pupille ronde. Les reptiles terrestres ont une pupille en fente verticale.

Les amphibiens ont des pupilles aux formes curieuses.
Chez les mammifères, seuls le chat, certains renards et le loir ont une pupille en fente verticale. Le cheval a une pupille en fente horizontale et, de surcroît, le bord libre supérieur possède une ou deux excroissances noires, les corpora nigra, ou grains de suie, auxquels on attribue le pouvoir de protéger la rétine des rayons venant du ciel.
B/L'acuité visuelle ou résolution
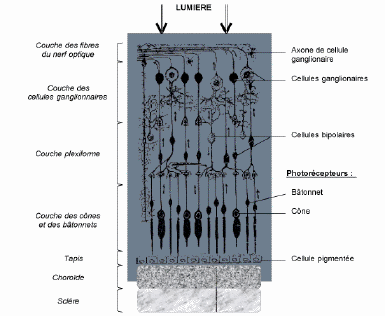
C'est la distance minimale entre deux points de l'objet susceptibles de donner une image distincte et nette. Elle dépend bien sûr de la qualité de la réfraction, mais aussi de la densité rétinienne en photorécepteurs et du nombre de cellules rétiniennes connectées à chaque fibre du nerf optique.
Chez les animaux diurnes, l'acuité dépendra aussi de l'intensité lumineuse stimulante, mettant en jeu, lorsqu'elle est élevée un plus grand nombre de cônes.
La richesse cellulaire de la rétine en photorécepteurs n'est pas la même selon la zone topographique. Chez les espèces dont la rétine possède à la fois cônes et bâtonnets, les cônes sont plus nombreux au centre qu'à la périphérie. On peut en outre mettre en évidence des régions spéciales
- l'area centralis,
- la macula,
- la fovea.
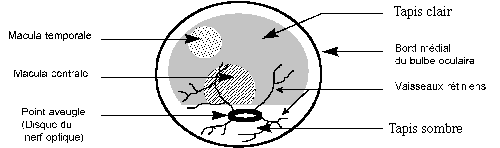
L'area centralis est une zone médio-temporale de plus grande richesse cellulaire. Elle correspond à un épaississement rétinien parfois macroscopiquement visible sous l'aspect d'une tache plus foncée.
Il existe une area chez les sélaciens, les amphibiens, quelques sauriens tel l'alligator et chez les carnivores, chiens et chats; chez les ruminants, elle a une disposition horizontale allongée qui la fait qualifier de striaeformis.
La macula est une zone avasculaire de plus grande richesse en cônes. Elle existe chez les primates, macroscopiquement visible à l'examen du fond d'oeil.
La fovea, uniquement constituée de cônes, présente en outre un amincissement dû à la raréfaction ou à la disparition de certaines couches rétiniennes .
Chez les téléostéens la fovea est rudimentaire et constituée de cellules amincies, sans raréfaction d'une couche rétinienne.
Chez les lacertiliens, existe tantôt une area tantôt une fovea, comme chez le caméléon.
Les oiseaux de basse-cour n'ont qu'une area, en revanche ; les rapaces ont deux foeva : une fovea centrale et une latérale. « Dans la fovea centrale, seule la couche des cellules ganglionnaires fait défaut celle des grains externes et internes s'amincissent seulement » (Rochon-Duvigneaud). Le bouquet central de cônes est beaucoup plus large, 1 500 cônes contre 200 chez l'homme.
L'acuité des oiseaux de proie est de loin la plus développée. Elle serait 7 à 8 fois supérieure à celle de l'homme.
C/La perception des couleurs
La perception des couleurs est liée à la présence dans la rétine de cônes sensibles au vert, au bleu et au jaune. L'étude de la vision des couleurs par l'animal fait appel à des techniques de modification comportementale et à des analyses spectrophotométriques de rayonnement absorbé par la rétine.
Sans pouvoir dire que l'unanimité soit faite, il semblerait qu'une majorité d'auteurs admettent les éléments suivants:
On ne sait pratiquement rien sur la vision des couleurs par les cyclostomes (lamproies).
La rétine des téléostéens perçoit la couleur : rouge, jaune, vert, bleu, violet et ultra-violet jusqu'à 365 nm.
Les amphibiens ont une bonne vision des couleurs, avec un maximum de perception pour le jaune en photopique et pour le vert en scotopique.
Chez les reptiles : la tortue différencie le bleu, le vert et l'orange ; le lézard différencie le jaune, le rouge, le vert et le bleu.
Les oiseaux ont un sens de la couleur très développé ; cela paraît logique eu égard à la richesse des coloris de leur plumage, laquelle n'a d'égale que la diversité chromatique des poissons.
L'oiseau règle son comportement sur la couleur beaucoup plus que sur la luminosité ou sur la forme.
La vision des couleurs par les mammifères est très discutée. Elle est certes associée au caractère diurne de l'animal. On admet que la musaraigne, l'écureuil sont parfaitement trichromates. Il semblerait que les bovins ne les distingueraient pas, ce qui ferait douter de l'efficacité de la muleta rouge au cours des corridas. Sur la capacité du chien, les avis sont très partagés ; quant au chat, on sait aujourd'hui qu'il est dichromate : l'étude histologique de sa rétine a montré qu'elle renfermait deux sortes de cônes, les uns sensibles au bleu indigo 450 nm, les autres à jaune-vert 556 nm. Dans les corps genouillés latéraux, certains neurones ont un maximum de sensibilité à 500 nm en basse luminance et 556 nm en forte luminance. Le chat ne peut donc distinguer en-dessous de 540 nm. Il lui est donc impossible de distinguer le rouge.
D/La perception du mouvement
Elle est très importante pour l'animal aussi bien pour l'exercice de la vigilance que pour la chasse d'une proie.
Elle dépend de deux facteurs:
- La qualité de la mosaïque rétinienne, c'est-à-dire la richesse de la rétine en photorécepteurs.
Quand les éléments rétiniens sont peu nombreux, la portion de rétine correspondant à une seule fibre du nerf optique est grande ; une image doit traverser une distance considérable avant d'exciter les éléments sensoriels associés à une autre fibre, de telle sorte qu'un petit mouvement risque de ne pas être apprécié.
- La persistance dans le temps de la vision.
En effet, si la stimulation persiste longtemps, un élément rétinien une fois stimulé ne peut réagir rapidement à un nouveau stimulus.
Le temps de persistance peut être étudié par la méthode du flicker avec enregistrement de l'E.R.G. La stimulation est faite à l'aide d'éclairs de plus en plus rapprochés jusqu'au moment où il n'y a plus de réponse rétinienne. La fréquence de stimula tion est dite alors fréquence de fusion ou F.F.F. (Flicker Fréquence de Fusion).
Chez la plupart des animaux, c'est surtout le haut niveau de la F.F.F. qui conditionne le haut degré de sensibilité au mouvement. les espèces qui se nourrissent de proies vivantes peuvent mourir de faim à côté de proies mortes. Les batraciens attrapent la nourriture qui bouge. De très nombreux animaux sauvages agissent exactement de la même façon.
E/La perception de l'espace
Elle consiste à apprécier l'espace et à l'intérieur de celui-ci la localisation des différents objets.
Cette perception peut se subdiviser en
- perception bidimensionnelle qui apprécie la position de l'objet, sa distance bidimensionnelle ;
-perception tridimensionnelle : perception de la profondeur qui exige une vision stéréoscopique binoculaire, extension de cette faculté à l'appréciation de la forme.
Dans cette perception de l'espace interviennent :
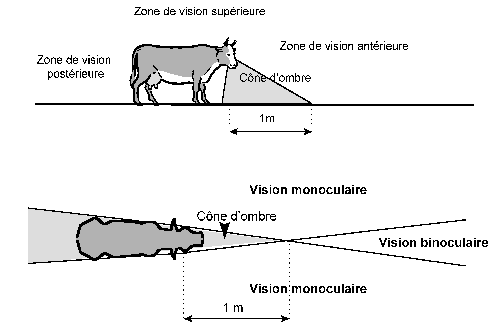
· le champ visuel monoculaire,
· le champ visuel binoculaire,
. les mouvements des globes oculaires,
. les mouvements de la tête.
Le champ visuel monoculaire varie peu chez les vertébrés. Sa valeur moyenne est de 170°. Il dépend essentiellement de trois facteurs :
- l'extension de la surface rétinienne
- l'incurvation de la surface admettant la lumière ;
- l'ouverture pupillaire effective.
D'après Duke-Elder, le large champ visuel du cheval est dû à l'extension de la rétine du côté nasal, alors que chez beaucoup d'oiseaux et chez les poissons de grande profondeur, la faible extension restreint ce même champ.
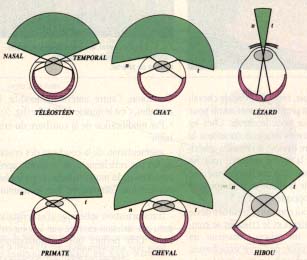
La cornée de l'homme représente un arc de 60 degrés et le champ est de 150 degrés ; chez le chat, la cornée est un arc de 170 degrés et le champ a 200 degrés.
L'ouverture pupillaire horizontale des équidés contribue également à élargir le champ visuel.
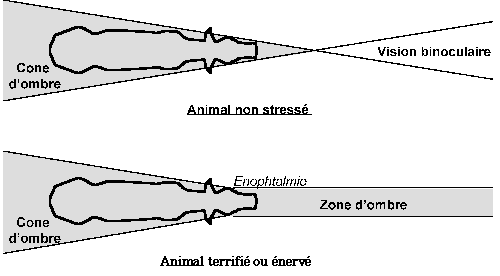
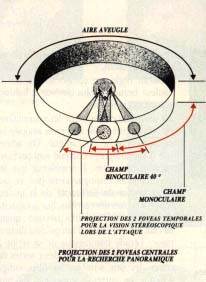
Le champ visuel binoculaire dépend essentiellement de la position des yeux:
-la position frontale élargit la vision binoculaire.
- la position latérale la restreint.

Les mouvements oculaires sont soit réflexes , soit volontaires et, dans ce dernier cas, les mouvements binoculaires peuvent être coordonnés ou non.
Chez tous les vertébrés autres que les mammifères, les mouvements oculaires volontaires sont toujours incoordonnés.
Enfin, il faut noter que chez les espèces à yeux peu mobiles, la flexuosité du cou compense très avantageusement ce handicap .
A l'issue de cette brève étude, on ne peut que souligner, malgré l'étendue des connaissances, le nombre des points qui demeurent obscurs. Pourquoi, peut-on répéter encore, tant de variété pour un même but ? La réponse est sans doute plus du domaine de la philosophie que de la médecine. Plus strictement scientifique est l'application à un plus grand nombre d'espèces des méthodes modernes d'investigation concernant en particulier la réfraction statique et la vision des couleurs.
Un secteur demeure cependant difficile à explorer à l'échelon individuel, c'est le champ visuel de l'animal vivant.
Parviendrons-nous unjour à faire une étude objective de ce champ ?
Il y a encore de beaux travaux à réaliser ; il est bon de ne pas en perdre la vision!